The Role of Losartan as a Potential Neuroregenerative Pharmacological Agent after Aneurysmal Subarachnoid Haemorrhage

 , and
, and
Abstract
:1. Introduction
2. Results
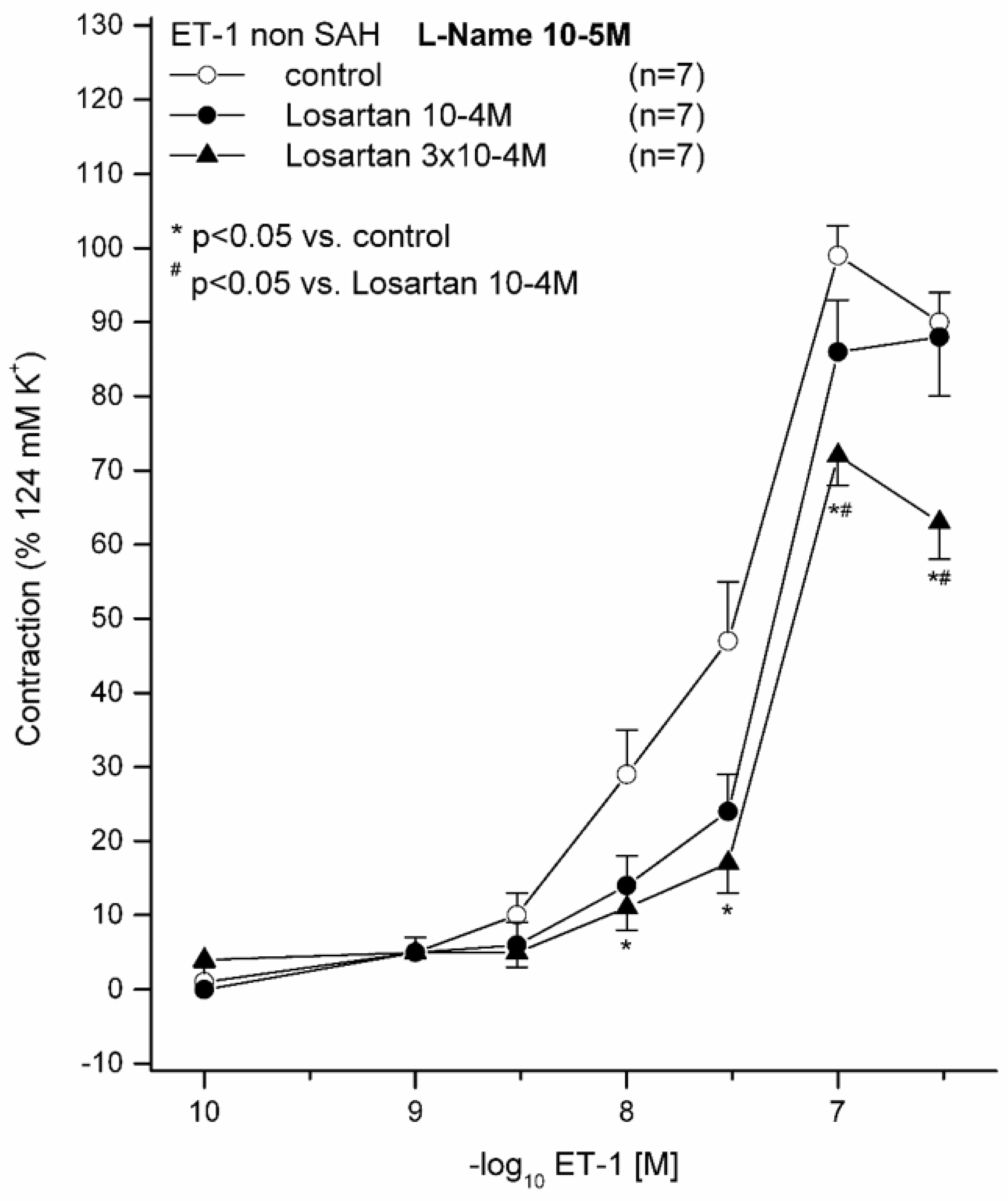
2.1. Effect of 10−5 M L-NAME on ET(A) Receptor-Induced Contraction in Sham Animals Preincubated with (10−4 M or 3 × 10−4 M) and without LOS
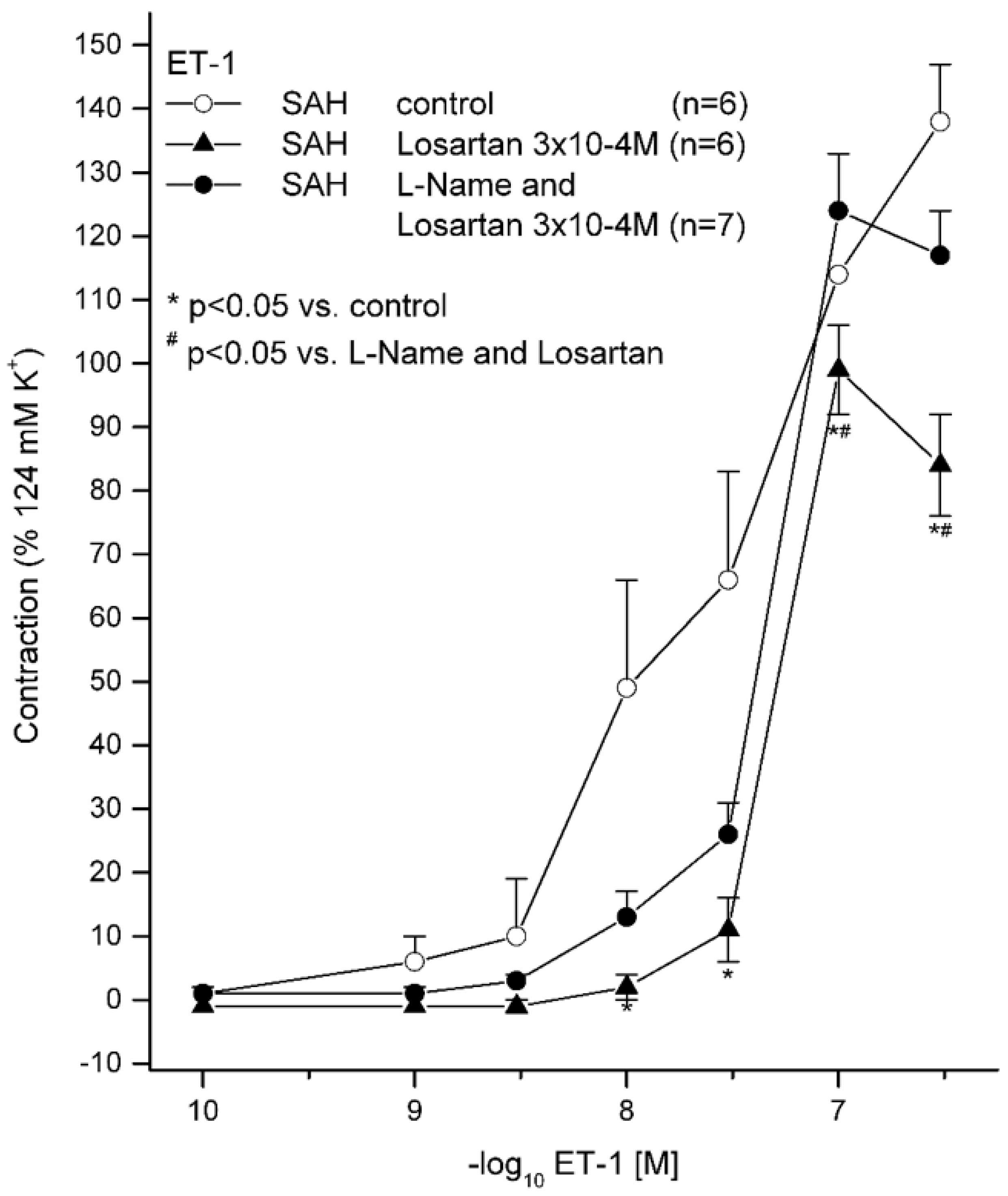
2.2. Effects of 10−5 M L-NAME with 3 × 10−4 M LOS on ET(A) Receptor-Induced Contraction and with (3 × 10−4 M) and without LOS on ET(A) Receptor-Induced Contraction in Subarachnoid Haemorrhage
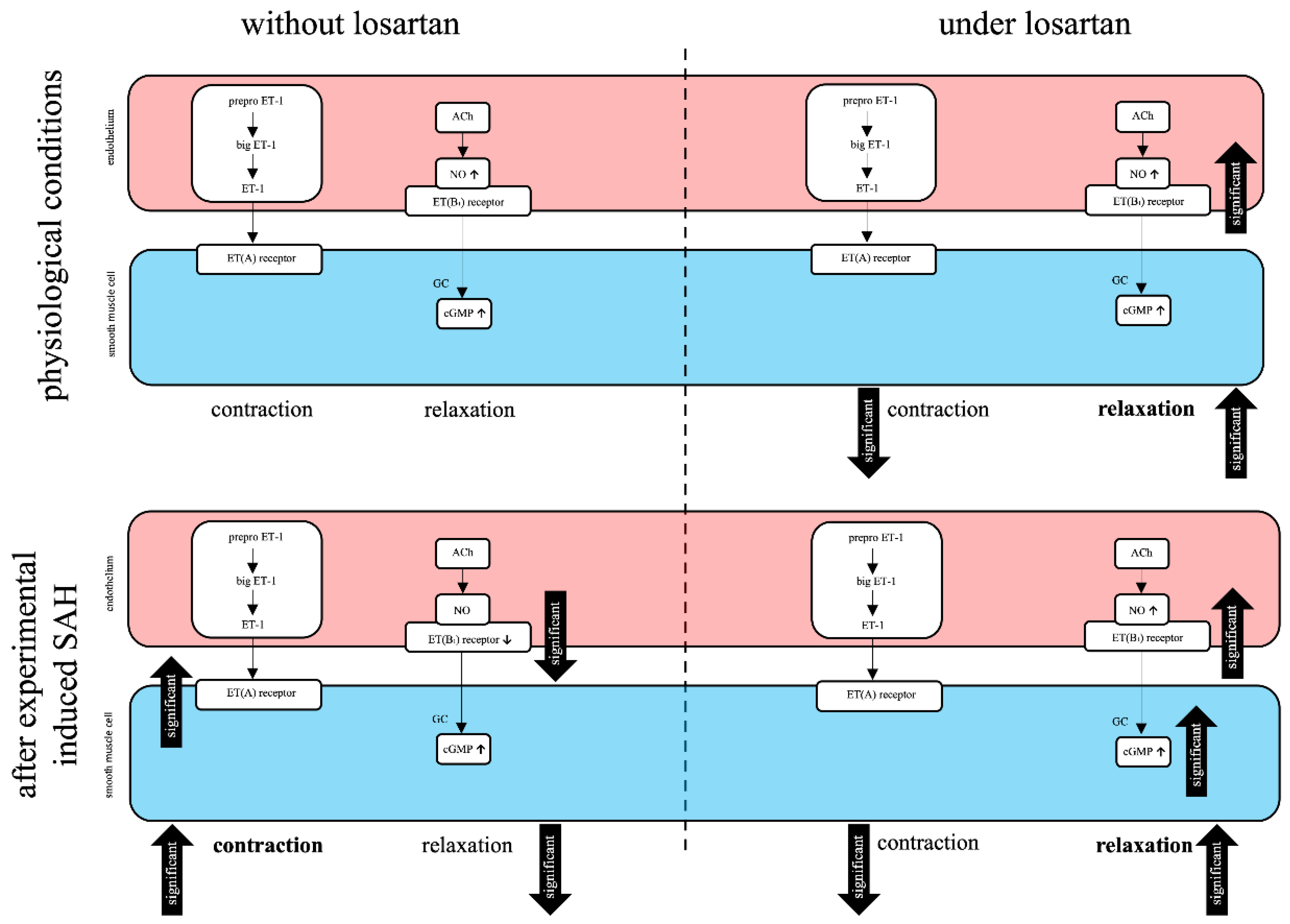
3. Discussion
3.1. The Role of LOS in a Translational Context
3.2. Further Perspectives
4. Material and Methods
4.1. Animals
4.2. Anaesthesia and Surgical Protocol
4.3. Further Processing of Brain Vessels
4.4. Experimental Setting
4.5. Compounds and Solvents
4.6. Analysis of Results and Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vergouwen, M.D.; Vermeulen, M.; van Gijn, J.; Rinkel, G.J.; Wijdicks, E.F.; Muizelaar, J.P.; Mendelow, A.D.; Juvela, S.; Yonas, H.; Terbrugge, K.G.; et al. Definition of delayed cerebral ischemia after aneurysmal subarachnoid hemorrhage as an outcome event in clinical trials and observational studies: Proposal of a multidisciplinary research group. Stroke 2010, 41, 2391–2395. [Google Scholar] [CrossRef] [Green Version]
- Salom, J.B.; Torregrosa, G.; Alborch, E. Endothelins and the cerebral circulation. Cereb. Brain Metab. Rev. 1995, 7, 131–152. [Google Scholar]
- Vatter, H.; Zimmermann, M.; Tesanovic, V.; Raabe, A.; Schilling, L.; Seifert, V. Cerebrovascular characterization of clazosentan, the first nonpeptide endothelin receptor antagonist clinically effective for the treatment of cerebral vasospasm. Part I: Inhibitory effect on endothelin(A) receptor-mediated contraction. J. Neurosurg. 2005, 102, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Andereggen, L.; Beck, J.; Z’Graggen, W.J.; Schroth, G.; Andres, R.H.; Murek, M.; Haenggi, M.; Reinert, M.; Raabe, A.; Gralla, J. Feasibility and Safety of Repeat Instant Endovascular Interventions in Patients with Refractory Cerebral Vasospasms. AJNR Am. J. Neuroradiol. 2017, 38, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Provencio, J.J. Inflammation in subarachnoid hemorrhage and delayed deterioration associated with vasospasm: A review. Acta Neurochir. Suppl. 2013, 115, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, M.; Yan, J.; Rolland, W.B.; Soejima, Y.; Caner, B.; Zhang, J.H. Early brain injury, an evolving frontier in subarachnoid hemorrhage research. Transl. Stroke Res. 2013, 4, 432–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreier, J.P.; Drenckhahn, C.; Woitzik, J.; Major, S.; Offenhauser, N.; Weber-Carstens, S.; Wolf, S.; Strong, A.J.; Vajkoczy, P.; Hartings, J.A.; et al. Spreading ischemia after aneurysmal subarachnoid hemorrhage. Acta Neurochir. Suppl. 2013, 115, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Budohoski, K.P.; Czosnyka, M.; Smielewski, P.; Kasprowicz, M.; Helmy, A.; Bulters, D.; Pickard, J.D.; Kirkpatrick, P.J. Impairment of cerebral autoregulation predicts delayed cerebral ischemia after subarachnoid hemorrhage: A prospective observational study. Stroke 2012, 43, 3230–3237. [Google Scholar] [CrossRef] [Green Version]
- Rubanyi, G.M.; Polokoff, M.A. Endothelins: Molecular biology, biochemistry, pharmacology, physiology, and pathophysiology. Pharmacol. Rev. 1994, 46, 325–415. [Google Scholar]
- Tirapelli, C.R.; Bonaventura, D.; Tirapelli, L.F.; de Oliveira, A.M. Mechanisms underlying the vascular actions of endothelin 1, angiotensin II and bradykinin in the rat carotid. Pharmacology 2009, 84, 111–126. [Google Scholar] [CrossRef]
- Konczalla, J.; Vatter, H.; Weidauer, S.; Raabe, A.; Seifert, V. Alteration of the cerebrovascular function of endothelin B receptor after subarachnoidal hemorrhage in the rat. Exp. Biol. Med. 2006, 231, 1064–1068. [Google Scholar]
- Ansar, S.; Vikman, P.; Nielsen, M.; Edvinsson, L. Cerebrovascular ETB, 5-HT1B, and AT1 receptor upregulation correlates with reduction in regional CBF after subarachnoid hemorrhage. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3750–H3758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saavedra, J.M.; Benicky, J.; Zhou, J. Mechanisms of the Anti-Ischemic Effect of Angiotensin II AT( 1 ) Receptor Antagonists in the Brain. Cell Mol. Neurobiol. 2006, 26, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Konczalla, J.; Wanderer, S.; Mrosek, J.; Schuss, P.; Platz, J.; Guresir, E.; Seifert, V.; Vatter, H. Crosstalk between the angiotensin and endothelin-system in the cerebrovasculature. Curr. Neurovasc. Res. 2013, 10, 335–345. [Google Scholar] [CrossRef]
- Hosmann, A.; Rauscher, S.; Wang, W.T.; Dodier, P.; Bavinzski, G.; Knosp, E.; Gruber, A. Intra-Arterial Papaverine-Hydrochloride and Transluminal Balloon Angioplasty for Neurointerventional Management of Delayed-Onset Post-Aneurysmal Subarachnoid Hemorrhage Vasospasm. World Neurosurg. 2018, 119, e301–e312. [Google Scholar] [CrossRef]
- Wanderer, S.; Mrosek, J.; Vatter, H.; Seifert, V.; Konczalla, J. Crosstalk between the angiotensin and endothelin system in the cerebrovasculature after experimental induced subarachnoid hemorrhage. Neurosurg. Rev. 2018, 41, 539–548. [Google Scholar] [CrossRef]
- Barhoumi, T.; Jallat, I.; Berthelot, A.; Laurant, P. Human recombinant erythropoietin alters the flow-dependent vasodilatation of in vitro perfused rat mesenteric arteries with unbalanced endothelial endothelin-1 / nitric oxide ratio. Can. J. Physiol. Pharmacol. 2011, 89, 435–443. [Google Scholar] [CrossRef]
- Wanderer, S.; Mrosek, J.; Gessler, F.; Seifert, V.; Konczalla, J. Vasomodulatory effects of the angiotensin II type 1 receptor antagonist losartan on experimentally induced cerebral vasospasm after subarachnoid haemorrhage. Acta Neurochir. 2018, 160, 277–284. [Google Scholar] [CrossRef]
- Tchekalarova, J.D.; Ivanova, N.M.; Pechlivanova, D.M.; Atanasova, D.; Lazarov, N.; Kortenska, L.; Mitreva, R.; Lozanov, V.; Stoynev, A. Antiepileptogenic and neuroprotective effects of losartan in kainate model of temporal lobe epilepsy. Pharmacol. Biochem. Behav. 2014, 127, 27–36. [Google Scholar] [CrossRef]
- Zhang, R.; Witkowski, S.; Fu, Q.; Claassen, J.A.; Levine, B.D. Cerebral hemodynamics after short- and long-term reduction in blood pressure in mild and moderate hypertension. Hypertension 2007, 49, 1149–1155. [Google Scholar] [CrossRef] [Green Version]
- Wanderer, S.; Gruter, B.E.; Strange, F.; Sivanrupan, S.; Di Santo, S.; Widmer, H.R.; Fandino, J.; Marbacher, S.; Andereggen, L. The Role of Sartans in the Treatment of Stroke and Subarachnoid Hemorrhage: A Narrative Review of Preclinical and Clinical Studies. Brain Sci. 2020, 10, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Zhang, Y.; Zhu, B.; Wu, C.; Chen, Y. Telmisartan regulates the development of cerebral ischemia by alleviating endoplasmic reticulum stress. Pharmazie 2018, 73, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Tchekalarova, J.D.; Ivanova, N.; Atanasova, D.; Pechlivanova, D.M.; Lazarov, N.; Kortenska, L.; Mitreva, R.; Lozanov, V.; Stoynev, A. Long-Term Treatment with Losartan Attenuates Seizure Activity and Neuronal Damage Without Affecting Behavioral Changes in a Model of Co-morbid Hypertension and Epilepsy. Cell Mol. Neurobiol. 2016, 36, 927–941. [Google Scholar] [CrossRef] [PubMed]
- van Gijn, J.; Kerr, R.S.; Rinkel, G.J. Subarachnoid haemorrhage. Lancet 2007, 369, 306–318. [Google Scholar] [CrossRef]
- Forder, J.P.; Munzenmaier, D.H.; Greene, A.S. Angiogenic protection from focal ischemia with angiotensin II type 1 receptor blockade in the rat. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H1989–H1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-Klein, G.; Cacheaux, L.P.; Kamintsky, L.; Prager, O.; Weissberg, I.; Schoknecht, K.; Cheng, P.; Kim, S.Y.; Wood, L.; Heinemann, U.; et al. Losartan prevents acquired epilepsy via TGF-beta signaling suppression. Ann. Neurol. 2014, 75, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Smeda, J.S.; Watson, D.; Stuckless, J.; Negandhi, A. Post-stroke losartan and captopril treatments arrest hemorrhagic expansion in SHRsp without lowering blood pressure. Vascul. Pharmacol. 2018, 111, 26–35. [Google Scholar] [CrossRef]
- Oku, N.; Kitagawa, K.; Imaizumi, M.; Takasawa, M.; Piao, R.; Kimura, Y.; Kajimoto, K.; Matsumoto, M.; Hori, M.; Hatazawa, J. Hemodynamic influences of losartan on the brain in hypertensive patients. Hypertens. Res. 2005, 28, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G.; NC3Rs Reporting Guidelines Working Group. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef]
- Vatter, H.; Weidauer, S.; Konczalla, J.; Dettmann, E.; Zimmermann, M.; Raabe, A.; Preibisch, C.; Zanella, F.E.; Seifert, V. Time course in the development of cerebral vasospasm after experimental subarachnoid hemorrhage: Clinical and neuroradiological assessment of the rat double hemorrhage model. Neurosurgery 2006, 58, 1190–1197. [Google Scholar] [CrossRef]
- Bederson, J.B.; Pitts, L.H.; Tsuji, M.; Nishimura, M.C.; Davis, R.L.; Bartkowski, H. Rat middle cerebral artery occlusion: Evaluation of the model and development of a neurologic examination. Stroke 1986, 17, 472–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatter, H.; Zimmermann, M.; Seifert, V.; Schilling, L. Experimental approaches to evaluate endothelin-A receptor antagonists. Methods Find Exp. Clin. Pharmacol. 2004, 26, 277–286. [Google Scholar] [PubMed]
- Gurzu, B.; Costuleanu, M.; Petrescu, G. Interactions between extracellular and intracellular administered angiotensin II in isolated rat portal vein rings. Rev. Med. Chir. Soc. Med. Nat. Iasi 2004, 108, 652–656. [Google Scholar] [PubMed]
- Vatter, H.; Konczalla, J.; Weidauer, S.; Preibisch, C.; Raabe, A.; Zimmermann, M.; Seifert, V. Characterization of the endothelin-B receptor expression and vasomotor function during experimental cerebral vasospasm. Neurosurgery 2007, 60, 1100–1109. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham (n = 21) | SAH (n = 19) | |
|---|---|---|
| Normal | 20 | 0 |
| Mild | 1 | 4 |
| Moderate | 0 | 8 |
| Severe | 0 | 7 |
| ET-1 | E(max) Contraction | pD2 | n | |
|---|---|---|---|---|
| Sham with 10−5 M L-NAME without LOS | 99 ± 11% | 7.57 ± 0.30 | 7 | |
| Sham with 10−5 M L-NAME and 10−4 M LOS | 91 ± 19% | 7.35 ± 0.08 | 7 | |
| Sham with 10−5 M L-NAME and 3 × 10−4 M LOS | 73 ± 12% * | 7.35 ± 0.08 | 7 |
| ET-1 | E(max) Relaxation | pD2 | n | |
|---|---|---|---|---|
| SAH without LOS | 139 ± 22% | 7.64 ± 0.52 | 6 | |
| SAH with LOS 3 × 10−4 M | 101 ± 12% * | 7.28 ± 0.06 | 6 | |
| SAH with L-NAME 10−5 M and LOS 3 × 10−4 M | 124 ± 24% | 7.33 ± 0.04 | 7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanderer, S.; Andereggen, L.; Mrosek, J.; Kashefiolasl, S.; Marbacher, S.; Konczalla, J. The Role of Losartan as a Potential Neuroregenerative Pharmacological Agent after Aneurysmal Subarachnoid Haemorrhage. Int. J. Mol. Sci. 2020, 21, 6496. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186496
Wanderer S, Andereggen L, Mrosek J, Kashefiolasl S, Marbacher S, Konczalla J. The Role of Losartan as a Potential Neuroregenerative Pharmacological Agent after Aneurysmal Subarachnoid Haemorrhage. International Journal of Molecular Sciences. 2020; 21(18):6496. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186496
Chicago/Turabian StyleWanderer, Stefan, Lukas Andereggen, Jan Mrosek, Sepide Kashefiolasl, Serge Marbacher, and Jürgen Konczalla. 2020. "The Role of Losartan as a Potential Neuroregenerative Pharmacological Agent after Aneurysmal Subarachnoid Haemorrhage" International Journal of Molecular Sciences 21, no. 18: 6496. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186496